Speciācija — sugu veidošanās: skaidrojums, izolācija un hibridizācija
Uzzini, kā rodas sugas: specācija, izolācijas mehānismi un hibridizācija, DNS pierādījumi, piemēri un mūsdienu skatījums uz sugu veidošanos.
Sugu veidošanās ir saistīta ar sugu rašanos un diferencēšanos. Tā ir galvenā evolūcijas bioloģijas daļa, kas skaidro, kā no vienas cilts vai populācijas laika gaitā var rasties divas vai vairāk atšķirīgas taksonomiskas vienības.
Darvins uzskatīja, ka daudzas sugas var izveidoties pakāpenisku pārveidošanos rezultātā — to sauca par anagenēzi (jeb „flotisko” evolūciju). 20. gadsimta lielākā daļa bioloģu pievērsās citam skaitam procesu, proti, biežākai jaunu sugu rašanās formai, kur populācija sadalās divās vai vairākās neatkarīgās līnijās — to sauc par kladoģenēzi. Tradicionāli tika uzskatīts, ka kladoģenēzi veicina izolācijas mehānismi, kas traucē vai novērš genofona apmaiņu starp populācijām.
Attēlu galerija
10 Attēli









Speciācijas veidi
Speciācija notiek dažādos veidos, atkarībā no telpiskajām, ekoloģiskajām un ģenētiskajām attiecībām starp populācijām:
- Alopatriskā speciācija — sugas rodas tad, kad populācijas tiek fizikāli sadalītas (piem., salas, kalnu grēdas, ledus atdalījumi), un ilgstoša ģenētiskā izolācija noved pie atšķirībām. Tas ir klasisks un bieži sastopams scenārijs.
- Peripatriskā speciācija — mazai izolētai populācijai (dibinātāja efekts) ātri sakrājas jauni aleli; var rasties jaunas sugas, īpaši, ja adaptācija notiek jaunos apstākļos.
- Parapatriskā speciācija — blakus esošām populācijām ar daļēju genofona apmaiņu var rasties diverģence, ja pastāv gradienti vai robežas vides apstākļos.
- Simpatriskā speciācija — sugas rodas vienā un tajā pašā ģeogrāfiskajā teritorijā bez pilnīgas fizikālas izolācijas; parasti tiek saistīta ar dramatiskām ekoloģiskām vai reproduktīvām pārmaiņām (piem., barošanās preferences, meitēšanas uzvedība, vai ģenētiskais poliploīdisms augiem).
Izolācijas mehānismi
Izolācijas mehānismi, kas ierobežo vai novērš krustošanos un ģenētisko apmaiņu, iedalās galvenokārt divos lielos tipos:
- Priekšzigotiskie (prezygotic) — pasākumi, kas neļauj gametām satiekoties vai neļauj apaugļošanai notikt. Tai skaitā:
- laika (temporal) izolācija — sugas vai populācijas vairojas dažādos laikos;
- habitatālā izolācija — tās izmanto atšķirīgus mikrohabitat;
- uzvedības (behavioral) izolācija — atšķirīga zvanīšana, dejas vai citas signalizācijas sistēmas;
- mehāniskā izolācija — reproduktīvie orgāni nav saderīgi;
- gamētu nesaderība — spermatozoīdi un olšūnas nespēj apvienoties.
- Pēczigotiskie (postzygotic) — notiek pēc apaugļošanās un ietver:
- hibrīdu neizdzīvotspēju (hybrid inviability) — embriji vai jaunie indivīdi neattīstās;
- hibrīdu neauglība (hybrid sterility), piemēram, zirgu un ēzeļa hibrīda muļļa neauglība;
- hibrīdu pazīmes, kas ar laiku samazina pēcnācēju izdzīvotspēju (hybrid breakdown).
Hibridizācija, introgressija un mūsdienu secinājumi
Nav šaubu, ka fiziskā atdalīšanās bieži veicina sugu veidošanos — to ilustrē daudzi piemēri — tomēr pēdējo desmitgažu ģenētiskie pētījumi maina vai precizē šo skatu. Analizējot dzīvu organismu DNS secību, ir atklāts, ka starp radniecīgām sugām bieži notiek hibridizācija. Hibridizācija nozīmē, ka krustošanās rezultātā tiek pārnesti gēni no vienas populācijas vai sugas uz citu — šis process tiek saukts par introgressiju.
Šī ģenētiskā apmaiņa nozīmē, ka reproduktīvā izolācija nav vienīgā sugas definīcija un ka sugu veidošanās var notikt pat tad, ja dažādas līnijas saglabā ierobežotu gēnu plūsmu. Ir vairāki svarīgi secinājumi no mūsdienu pētījumiem:
- Speciācija var notikt ar daļēju genofona apmaiņu (speciācija ar gēnu plūsmu). Dabiska izvēle var nostiprināt atšķirības tajās genomikas daļās, kas ietekmē reproduktīvos vai ekoloģiskos uztveres signālus.
- Hibridizācija dažkārt noved pie jaunas sugas rašanās (hibrīdu sugu rašanās), īpaši augos, kur poliploīdija (hromosomu daudzuma dubultošanās) var ātri radīt reproduktīvu izolāciju. Piemēri: daudzas jaunas sugu grupas, piemēram, dažas augu sugas (Tragopogon u.c.), radušās hibridizācijas un poliploīdijas rezultātā.
- Hibridzonas (teritorijas, kur populācijas krustojas) var būt ilgstoši stabilas un kalpo kā laboratorijas dabā, kur novērot adaptācijas, selekciju pret hibrīdiem vai pretēji — selekciju, kas veicina hibrīdu panākumus.
- Genoma līmenī var rasties „diferencēšanas saliņas” (islands of divergence) — ģenoma reģioni, kurus spēcīgi ietekmē selekcija, kamēr pārējā genomā notiek gēnu apmaiņa. Tas skaidro, kā diverģence var pastāvēt selektīviem lokusiem, pat ja pārējais genoms tiek homogenizēts.
Papildus jēdzieni un piemēri
Vairāki labi zināmi piemēri palīdz ilustrēt speciācijas mehānismus:
- Darvina dzeņu (Galapagosa dzeņu) variācijas — ekoloģiskā izolācija un dabiskā izvēle ieguvušas plašu uzmanību kā klasiskā adaptīvā radiācija piemērs.
- Ensatina salamandu „gredzena sugas” piemērs — pakāpeniska ģeogrāfiska variācija, kas beidzot rada divas neatgriezeniski atšķirīgas formas, kas vietās nesapārojas.
- Augos — bieži tiek novērota hibrīdu sugu rašanās, īpaši, ja poliploīdija nodrošina tūlītēju reproduktīvu izolāciju.
- Dzīvniekos — hibridizācija un genoma introgressija ir konstatēta, piemēram, starp polārlāčiem un brūnajiem lāčiem, kā arī tauriņiem (Heliconius) un zivīm (cichlidi), kur gēnu apmaiņa ietekmē krāsojumu vai barošanās īpašības.
Nobeigums
Speciācija nav vienkāršs un lineārs process — tā ietver mijiedarbību starp ģeogrāfiju, ekoloģiju, uzvedību, ģenētiku un nejaušiem notikumiem. Lai gan fizikāla atdalīšanās (alopatrija) bieži ir izšķiroša, mūsdienu ģenētiskie pētījumi parāda, ka hibridizācija, introgressija un selektīva atšķirība genomā var radīt jaunas sugas arī bez pilnīgas izolācijas. Tādējādi sugu definīcija un speciācijas mehānismi ir dinamiski jēdzieni, kas jātver kā komplekss procesu tīklojums, nevis kā viena noteikta ceļa rezultāts.
Izolācijas mehānismi
Izolācijas mehānismi ir tādi, kas kavē veiksmīgu vairošanos starp sugas grupām. Ir izveidota populāciju reproduktīvā izolācija. Tas ir īpaši svarīgi bioloģiskās sugas jēdzienam, jo sugas tiek definētas pēc reproduktīvās izolācijas.
Izolācijas mehānismus var iedalīt divās grupās - pirms un pēc apaugļošanas.
Pirms apaugļošanas
Faktori, kas neļauj indivīdiem pāroties.
- Ģeogrāfiskā izolācija: Sugas sastopamas dažādās teritorijās, un bieži vien tās atdala barjeras.
- Izolācija laikā: Indivīdi nesadraudzējas, jo tie ir aktīvi dažādos laikos. Tas var būt dažādos diennakts laikos vai dažādos gadalaikos. Sugu pārošanās periodi var nesakrist. Indivīdi nesatiek viens otru vai nu pārošanās periodos, vai vispār nesatiekas.
- Ekoloģiskā izolācija: Indivīdi pārojas tikai sev vēlamajā biotopā. Tie nesaskaras ar citu sugu īpatņiem, kam ir atšķirīgas ekoloģiskās preferences.
- Uzvedības izolācija: Dažādu sugu indivīdi var satikties, bet viens no tiem neatpazīst nekādus seksuālus signālus, kas var tikt doti. Indivīds vairumā gadījumu izvēlas savas sugas pārstāvi.
- Mehāniskā izolācija: var mēģināt kopulāciju, bet spermas pārnese nenotiek. Indivīdi var būt nesaderīgi lieluma vai morfoloģijas dēļ.
- Gametic nesaderība: Spermas pārnese notiek, bet olšūna netiek apaugļota.
Pēc apaugļošanas
Faktori, kas kavē veiksmīgu pārošanos, piemēram, ģenētiskā nesaderība, hibrīda neauglība vai sterilitāte.
- Zigotiskā mirstība: Olšūna tiek apaugļota, bet zigota neattīstās.
- Hibrīda dzīvotnespēja: veidojas hibrīda embrijs, bet tas nav dzīvotspējīgs.
- Hibrīda sterilitāte: hibrīds ir dzīvotspējīgs, bet iegūtais pieaugušais īpatnis ir sterils.
- Hibrīda sadalījums: Pirmās paaudzes (F1) hibrīdi ir dzīvotspējīgi un auglīgi, bet nākamās hibrīdu paaudzes (F2 un atpakaļ krustojumi) ir dzīvotnespējīgas vai neauglīgas.

Ģeogrāfiskā izolācija
Tiek uzskatīts, ka tas ir visbiežāk sastopamais sugu veidošanās iemesls. Pirmais par to domāja Morics Vāgners, vācu pētnieks un dabas vēsturnieks.
Vāgnera karjeras sākums bija ģeogrāfa karjera, un viņš publicēja vairākas ģeogrāfiskas grāmatas par Ziemeļāfriku, Tuvajiem Austrumiem un Tropu Ameriku. Viņš bija arī aizrautīgs dabaszinātnieks un kolekcionārs, un tieši ar šo darbu viņš ir vislabāk pazīstams biologu vidū. Ernsts Mairs, evolūcijas pētnieks un bioloģijas vēsturnieks, ir aprakstījis Vāgnera nozīmi. 562.-565. lpp.
Trīs gadu laikā, ko Vāgners pavadīja Alžīrijā, viņš cita starpā pētīja nelidojošās vaboles Pimelia un Melasoma. Katra ģints ir sadalīta vairākās sugās, un katra no tām ir ierobežota ar kādu ziemeļu piekrastes posmu starp upēm, kas no Atlasa kalniem nolaižas līdz Vidusjūrai. Tiklīdz tiek šķērsota kāda upe, parādās cita, bet tuvu radniecīga suga.
"... sākusies suga [radīsies] tikai tad, kad daži indivīdi pārsniegs savas izplatības robežas... jaunas rases veidošanās nekad neizdosies... bez ilgstošas kolonistu nošķirtības no citiem savas sugas pārstāvjiem".
Tas bija agrīns viena veida ģeogrāfiskās sugu veidošanās procesa apraksts. To 1942. gadā no jauna ieviesa Mairs, un ģeogrāfiskās specializācijas nozīme kļuva par vienu no evolūcijas sintēzes pamatidejām.
Cits ģeogrāfiskās sugas veidošanās apzīmējums ir alopatriskā sugas veidošanās. Allopatrija nozīmē "atšķirīga zeme".
Brīvas salas
Vulkāniskās salas veidojas bez dzīvības, un visa dzīvība ir jānogādā ar vēja vai ūdens palīdzību. No Havaju salām un Galapagu salām mēs zinām, ka visas dzīvības formas mainās, kad tās sasniedz salas no kontinenta.
Havaju salās, kuru platība ir aptuveni 6500 kvadrātkilometru (17 000 km2), ir visdaudzveidīgākā drozofīlo mušu kolekcija pasaulē, kas dzīvo gan lietus mežos, gan kalnu pļavās. Ir zināmas aptuveni 800 Havaju salu drozofilu sugas.
Pētījumi liecina par skaidru sugu "plūsmu" no vecākām uz jaunākām salām. Ir arī gadījumi, kad sugas kolonizējas atpakaļ uz vecākām salām un izlaiž salas, bet tie ir daudz retāki.
Saskaņā ar kālija/argoņa radioaktīvo datēšanu pašreizējās salas datētas no 0,4 miljoniem gadu (mya) (Mauna Kea) līdz 10mya (Necker). Vecākais Havaju arhipelāga loceklis, kas joprojām atrodas virs jūras, ir Kure atols, kas datējams ar 30 mya.
Pats arhipelāgs, kas izveidojies Klusā okeāna plātnei pārvietojoties pāri karstajam punktam, ir pastāvējis daudz ilgāk, vismaz līdz krīta laikam. Havaju salas un bijušās salas, kas tagad atrodas zem jūras, veido Havaju un Imperatora jūras kalnu ķēdi, un daudzi zemūdens kalni ir gajoti.
Acīmredzot visas Havaju salās sastopamās drozofīlu sugas ir cēlušās no vienas senču sugas, kas salas kolonizēja pirms aptuveni 20 miljoniem gadu. Turpmāko adaptīvo radiāciju veicināja konkurences trūkums un plašais brīvo nišu klāsts. Lai gan būtu iespējams, ka salu kolonizēja viena grūsna mātīte, tomēr daudz ticamāk, ka tā ir bijusi vienas sugas dzīvnieku grupa.
Havaju salu arhipelāgā ir arī citi dzīvnieki un augi, kas piedzīvojuši līdzīgu, lai gan ne tik iespaidīgu adaptācijas radiāciju.
Gredzenu sugas
Bioloģijā gredzenveida suga ir savienota virkne blakusesošu populāciju, no kurām katra var krustoties ar blakusesošajām populācijām. Abi ķēdes gali pārklājas.
Abas gala populācijas sērijā ir pārāk tālu radniecīgas, lai krustotos. Šādas ģenētiski saistītas "gala" populācijas, kas nav radniecīgas, var līdzāspastāvēt vienā un tajā pašā reģionā, tādējādi noslēdzot "gredzenu".
Gredzenveida sugas ir svarīgs evolūcijas pierādījums: tās parāda, kas laika gaitā notiek, populācijām ģenētiski atšķiroties. Ričards Dorkinss (Richard Dawkins) novēroja, ka gredzenveida sugas "tikai telpiskajā dimensijā mums parāda kaut ko tādu, kam vienmēr jānotiek laika dimensijā".
Tomēr ir grūti atrast vienkāršu un vienkāršu piemēru.
Larus kaijas
Klasisks gredzenveida sugas piemērs ir Larus kaiju cirkumpolārais sugas gredzens. Šo kaiju areāls veido gredzenu ap Ziemeļpolu, ko parasti atsevišķas kaijas nepārlido.
Vispārīgi runājot, kaijas var zināmā mērā krustoties ar savām kaimiņvalstīm, izņemot abos gredzena galos.
Mazās melnkaklajās kaijas un reņģes kaijas ir pietiekami atšķirīgas, lai tās parasti nekrustotos; tādējādi kaiju grupa veido nepārtrauktu grupu, izņemot vietas, kur abas līnijas sastopas Eiropā.
Lai gan realitāte ir daudz sarežģītāka, tā labi parāda vispārējo ideju.
Ensatina salamandras
Ensatina salamandrs ir gredzenveida suga kalnos ap Kalifornijas Centrālo ieleju. Komplekss veido pakava formu ap kalniem. Lai gan katra no 19 populācijām var krustoties starp 19 populācijām, kas atrodas ap zemakša, Ensatina eschscholtzii pasuga, kas atrodas zemakša rietumu galā, nevar krustoties ar Ensatina klauberi, kas atrodas austrumu galā. Tas ilustrē "gandrīz visus sugu veidošanās procesa posmus" (Dobžanskis). Ričards Hightons apgalvoja, ka Ensatina ir vairāku sugu gadījums, nevis vienas sugas kontinuitāte.
Zaļganā dzilnīte
Zaļganās dzilnītes (Phylloscopus trochiloides) ir vairākas pasugas, no kurām Eiropā pazīstamākā ir P. t. viridianus. Tā ir gredzenveida suga, kuras populācijas atšķiras uz austrumiem un rietumiem no Tibetas plakankalnes, bet vēlāk satiekas ziemeļu pusē. To savstarpējās attiecības ir diezgan neskaidras.




Simpatriskā sugu veidošanās
Simpatriskā sugu veidošanās ir divu vai vairāku sugu pēcnācēju veidošanās no vienas senču sugas, kas dzīvo vienā un tajā pašā ģeogrāfiskajā vietā. Mūsdienās tiek uzskatīts, ka tas ir ļoti bieži sastopams.
Simpatriskās sugu sugas atšķiras, dzīvojot vienā un tajā pašā vietā. Bieži minēti simpatriskās sugu sastopamības piemēri ir kukaiņi, kas kļūst atkarīgi no dažādiem saimniekaugiem vienā un tajā pašā teritorijā.
Simpatriskās sugu veidošanās kā sugu veidošanās mehānisma pastāvēšana tika karsti apstrīdēta. Cilvēki iebilda, ka simpātiskās sugas veidošanās pierādījumi patiesībā ir mikroģeogrāfiskās sugas veidošanās piemēri. Kopumā tagad šāds skaidrojums vairs netiek uzskatīts par vēlamo. Drīzāk tā ir radniecīgu sugu krustošanās rezultāts, kam seko dabiskā atlase, kas iedarbojas uz šādu krustojumu pēcnācējiem. Viens no plaši atzītiem simpātiskas sugu veidošanās piemēriem ir Nabugabo ezera cichlidu suga Austrumāfrikā, ko, kā uzskata, izraisīja dzimumselekcija.
Speciācija poliploidizācijas ceļā
Poliploīdija ir izraisījusi daudzus straujus sugu veidošanās gadījumus, jo, piemēram, tetraploīdu x diploīdu pārošanās pēcnācēji bieži iegūst triploīdus sterilus pēcnācējus.
Tomēr ne visi poliploīdi ir reproduktīvi izolēti no saviem vecāku augiem, un gēnu plūsma joprojām var notikt, piemēram, izmantojot triploīdu hibrīdu x diploīdu pārošanos, kas rada tetraploīdus.
Daudzas no esošajām augu un lielākā daļa dzīvnieku sugu evolūcijas vēsturē acīmredzot ir piedzīvojušas poliploidizāciju. Veiksmīgu poliploīdu sugu vairošanās dažkārt ir aseksuāla, ar partenogēzi. Nezināmu iemeslu dēļ daudzi aseksuāli organismi ir poliploīdi.
Gaiļdegleņu muša
Viens no evolūcijas darba piemēriem ir gadījums ar vilkābeles mušu Rhagoletis pomonella, kas, šķiet, piedzīvo simpatriju sugu veidošanos.
Dažādas vilkābeles populācijas barojas ar dažādiem augļiem. Atsevišķa populācija parādījās Ziemeļamerikā 19. gadsimtā kādu laiku pēc tam, kad tika introducētas ābeles, kas nav vietējās sugas. Šī ar āboliem barojošā populācija parasti barojas tikai ar āboliem, nevis ar vēsturiski iecienītajiem vilkābeles augļiem. Pašreizējā populācija, kas barojas ar vilkābeles augļiem, parasti neēd ābolus.
Daži pierādījumi liecina, ka notiek simpātiska sugu veidošanās. Gailenes mušas nobriest vēlāk sezonā un tām ir nepieciešams ilgāks laiks, lai nobriestu, nekā ābeļu mušām, un ir maz pierādījumu par krustošanos (pētnieki ir dokumentējuši 4-6 % hibridizācijas līmeni).
Jaunās vilkābeles mušas parādīšanās ir progresējošas evolūcijas piemērs.
Hibridizācija
Retos gadījumos jauna suga veidojas, kad pārojas dažādu sugu pārstāvji. Parasti šādu hibrīdu pārošanās produkti ir neauglīgi (neauglīgi) vai relatīvi neauglīgi, un tāpēc dabiskā atlase tos iznīcina.
Ir atrasts viens jaunas un veiksmīgas hibrīda sugas piemērs. Jaunā suga ir itāļu un spāņu zvirbuļu hibrīds, un pamatfakti ir pārbaudīti, veicot DNS sekvenču analīzi asinīs. Šis putns dzīvo Itālijā, reģionā, kur dzīvo abas vecāku sugas. Tas nevairojas kopā ar spāņu zvirbuļiem, lai gan dzīvo ar tiem līdzās.
Mākslīga sugu veidošanās
Jaunas sugas ir radītas, izmantojot pieradināto dzīvnieku audzēšanu, taču sākotnējie datumi un metodes nav skaidri zināmi. Piemēram, mājas aitas tika radītas hibridizācijas ceļā, un tās vairs nespēj radīt dzīvotspējīgus pēcnācējus ar Ovis orientalis - sugu, no kuras tās ir cēlušās.
Savukārt mājas liellopus var uzskatīt par vienu sugu ar vairākām savvaļas vēršu, gauru, jaku u. c. šķirnēm, jo tie viegli iegūst auglīgus pēcnācējus.
Laboratorijas sugas
Vislabāk dokumentētie jaunu sugu radīšanas gadījumi laboratorijā tika veikti 20. gadsimta 80. gadu beigās. Viljams Raiss un G. V. Sāls (William Rice and G. W. Salt) izaudzēja augļmutes Drosophila melanogaster, izmantojot labirintu ar trim dažādām dzīvotņu izvēlēm, piemēram, gaišs/tumss un mitrs/sauss.
Katra paaudze tika ievietota labirintā, un mušu grupas, kas iznāca no divām no astoņām izejām, tika nodalītas, lai vairotos savā grupā. Pēc trīsdesmit piecām paaudzēm abas grupas un to pēcnācēji tika reproduktīvi izolēti, jo tām bija izteiktas biotopa preferences: tās pārojās tikai tajās zonās, kurām tās deva priekšroku, un tādējādi nepārokojās ar mušām, kas deva priekšroku citām zonām. Šādu mēģinājumu vēsture ir aprakstīta Rice un Hostert (1993).
Diāna Dodda arī spēja parādīt, kā Drosophila pseudoobscura reproduktīvā izolācija var attīstīties no pārošanās preferencēm tikai pēc astoņām paaudzēm, izmantojot dažādus barības veidus - cieti un maltozi.
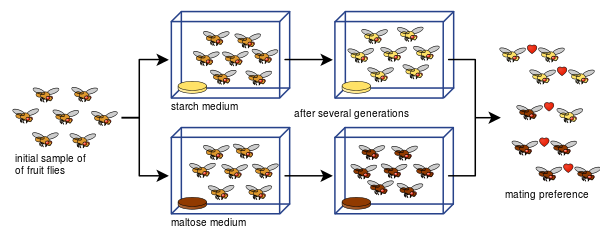
Dodda eksperimentu daudzi citi ir viegli atkārtojuši, tostarp ar cita veida augļmušām un pārtikas produktiem.
Pastiprinājums (Wallace efekts)
Pastiprināšana ir process, kurā dabiskā atlase palielina reproduktīvo izolētību.
Tas var notikt pēc tam, kad divas vienas sugas populācijas ir atdalītas un pēc tam atkal nonāk saskarē. Ja to reproduktīvā izolācija bija pilnīga, tad tās jau būs izveidojušās par divām atsevišķām nesaderīgām sugām.
Ja to reproduktīvā izolācija ir nepilnīga, tad turpmākas pārošanās starp populācijām radīs hibrīdus, kas var būt vai nebūt auglīgi. Ja hibrīdi ir neauglīgi vai auglīgi, bet mazāk piemēroti nekā viņu priekšteči, tad turpmāka reproduktīvā izolācija vairs nenotiks un sugas veidošanās būtībā ir notikusi (piemēram, kā zirgu un ēzeļu gadījumā).
Tas tiek pamatots ar to, ka, ja hibrīda pēcnācēju vecākiem katram ir dabiski selekcionētas īpašības, kas raksturīgas atšķirīgai videi, hibrīda pēcnācējam būs abu vecāku īpašības, un tas neatbildīs nevienai no šīm nišām tik labi kā viens no vecākiem. Hibrīdu zemā fitnesa dēļ selekcija dotu priekšroku asortatīvai pārošanai, kas samazinātu hibridizāciju.
Dažkārt to dēvē par Vollesa efektu pēc evolūcijas biologa Alfrēda Rasela Vollesa (Alfred Russel Wallace) vārda, kurš 19. gadsimta beigās ierosināja, ka tas varētu būt svarīgs faktors sugu veidošanās procesā.
Ja hibrīdu pēcnācēji ir piemērotāki par saviem priekštečiem, tad populācijas atkal saplūdīs vienā sugā tajā teritorijā, kurā tās saskaras.
Ja starp divām sugas formām ir "hibrīda zona", atdalīšanai ir nepieciešama pastiprināšana. Hibrīdu zonas ir reģioni, kuros sastopas un krustojas atšķirīgas populācijas. Šajos reģionos ļoti bieži sastopami hibrīdi pēcnācēji, kas parasti rodas, atšķirīgām sugām nonākot sekundārā kontaktā.
Bez pastiprināšanas abas populācijas vai sugas nekontrolēti krustotos. Pastiprināšanu var izraisīt mākslīgās atlases eksperimentos, kā aprakstīts iepriekš.
Jautājumi un atbildes
J: Kas ir speciācija?
A: Sugu veidošanās ir process, kā veidojas sugas. Tā ir galvenā evolūcijas bioloģijas daļa.
J: Ko Darvins domāja par sugu veidošanos?
A: Darvins uzskatīja, ka lielākā daļa sugu radās tieši no jau eksistējošām sugām, ko viņš sauca par anagenēzi jeb "filoģenētisko evolūciju".
J: Kā sugu veidošanos kopumā vērtēja 20. gadsimtā?
A.: 20. gadsimtā zinātnieki uzskatīja, ka lielākā daļa sugu veidojas, sadaloties agrākām sugām, ko dēvēja par kladoģenēzi. Tika arī uzskatīts, ka šo sadalīšanos izraisīja vai veicināja tādi izolācijas mehānismi kā fiziska atdalīšanās.
J: Kā jaunākie pētījumi ir mainījuši mūsu izpratni par sugu veidošanos?
A: Nesenie pētījumi ir pierādījuši, ka var notikt radniecīgu sugu krustošanās, un šādā krustošanās procesā var tikt nodoti gēni. Tas nozīmē, ka sugas noteikšanai ne vienmēr ir nepieciešama reproduktīvā izolācija, un sugas veidošanās procesā var nebūt nepieciešama alopatrija (fiziska atdalīšana).
J: Kāda saistība fiziskai nošķiršanai ir ar jaunu sugu veidošanos?
A: Tiek uzskatīts, ka fiziska atdalīšanās starp kādreiz līdzās dzīvojošām sugām ir galvenais faktors jaunu sugu veidošanās procesā, ko ilustrē daudzi zinātniskajā literatūrā aplūkotie piemēri.
Saistītie raksti
Autors
AlegsaOnline.com Speciācija — sugu veidošanās: skaidrojums, izolācija un hibridizācija Leandro Alegsa
URL: https://lv.alegsaonline.com/art/92570
Avoti
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833